I rettili sono molto curiosi e offrono, fra passato e presente, una vastissima gamma di comportamenti e soluzioni evolutive, in una enorme quantità di ambienti (acqua, aria con gli uccelli e gli pterosauri e terra, dai deserti alle foreste pluviali, solo le aree polari sono precluse) e di riproduzione (ovipari, vivipari, ovovivipari). Anche il movimento non fa eccezione: nella loro storia hanno alternato una andatura che striscia sul suolo con arti corti (o addirittura senza), quadrupede con arti di discreta lunghezza, fino a bipedi obbligati in forme provviste di lunghi arti posteriori, mentre in mare dalle tartarughe agli ittiosauri arti e/o coda sono stati usati in vari modi per nuotare. Oggi i bipedi per eccellenza fra i discendenti dei rettili triassici sono gli uccelli, ma anche qualche lucertola è capaci di muoversi su due zampe per alcuni tratti. Mi pareva che anche il Tuatara si comporti così, ma non ho trovato conferme. Quindi fra i diapsidi (rettili e uccelli) l’acquisizione di una andatura bipede facoltativa insieme o alternativa a quella strisciante sembra essere avvenuto svariate volte in svariati gruppi. Invece nei mammiferi se si escludono i canguri, un bipede obbligato come Homo e la possibilità di stazione eretta per le scimmie antropomorfe, il bipedismo è estremamente raro. Il perché di questa differenza comportamentale non è ancora stato ben determinato, ma una nuova ricerca suggerisce una soluzione piuttosto intelligente nelle differenze fra mammiferi e rettili nella anatomia della parte posteriore del bacino.

I rettili si dividono grossolanamente in Arcosauri (uccelli, coccodrilli e dinosauri) e Lepidosauri (lucertole, serpenti, varani etc). La collocazione delle tartarughe e degli altri rettili marini mesozoici, è più discussa. Adesso sta prevalendo l’idea che si tratti di arcosauri molto basali. Ne avevo parlato qui a proposito delle tartarughe e qui a proposito degli altri rettili marini.
Il fossile più antico mai ritrovato che mostra l’evidenza della capacità di una andatura bipede in un rettile è Eudibamus cursoris, un piccolo Bolosauro trovato in Germania [1]. Siamo nel Permiano inferiore (290 milioni di anni fa) quindi addirittura in tempi ben precedenti alla divisione fra lepidosauri e arcosauri, che dovrebbe essere avvenuta nel Permiano superiore: l’ultimo lavoro in merito parla di almeno 260 milioni di anni fa [2]; diciamo quindi che questa divergenza dovrebbe essere avvenuta addirittura prima della estinzione di massa della fine del Permiano medio (Guadalupiano), la quale a sua volta si colloca circa 8 milioni di anni prima della tragedia di fine Permiano.
La comparsa negli arcosauri dell’andatura bipede è successiva alla divergenza fra gli antenati dei coccodrilli (Pseudosuchia) e di quelli dei dinosauri (Avemetatarsalia). Questa divergenza ha coinvolto cambiamenti molto importanti nella forma e dimensioni del corpo e specialmente degli arti. Le vestigia più antiche di dinosauromorfi sono impronte lasciate da creature quadrupedi di circa 250 milioni di anni fa, di poco successive alla madre di tutte le estinzioni, quella di fine Permiano [3]. È evidente che all’inizio del Triassico i pochi superstiti, avendo campo libero, abbiano compiuto delle grandi radiazioni evolutive e abbiano avuto la possibilità di differenziarsi in maniera piuttosto veloce.
L’andatura bipede è, comunque, un tratto basale dei primi dinosauri triassici e dei loro parenti più stretti, ma nel Triassico - quindi prima dell’età dei dinosauri propriamente detta - questa caratteristica era diffusa anche in altri arcosauri prima che l’estinzione di fine Triassico segnasse la fine di tutte queste linee a parte dinosauri e rettili volanti. Insomma, sembra che all’epoca ci sia stata una diffusa pressione selettiva a favore di questa andatura negli arcosauri terrestri.
 |
| Ricostruzione di Teleocrater rhadinius, un antenato quadrupede dei dinosauri recentemente scoperto, da [4] |
Recentemente è stato pubblicato un articolo in cui viene descritto Teleocrater rhadinus [4], un quadrupede vissuto nel Triassico medio che accanto ad aspetti morfologici degli antenati comuni di coccodrilli e dinosauri, mostra anche delle caratteristiche tipiche di questi ultimi. In particolare quello che colpisce del fossile sono le zampe particolarmente lunghe per un rettile, come ne sono dotati solo i dinosauri attuali (gli uccelli).
 |
| Il Basilisco che corre sull'acqua con andatura bipede |
RETTILI BIPEDI ATTUALI AL DI FUORI DEGLI UCCELLI. Le poche specie di rettili attuali che esibiscono una andatura bipede (una cinquantina appartenenti a più gruppi di lucertole, in particolare iguanidi) per lo più sono dotate di forme snelle e arti più lunghi della media e si muovono così solo in particolari condizioni e per tempi molto brevi, soprattutto quando hanno bisogno di correre. Mentre queste lucertole corrono su due zampe il tronco si solleva e gli arti anteriori lasciano il terreno. Famoso al proposito è il Basilisco, che corre su due zampe sulla superficie dell’acqua. Non sono quindi bipedi “obbligati” e neanche “preferenziali”, al contrario degli antenati dei dinosauri, i quali, comunque, avevano gli arti ben più lunghi di quelli del rettile “medio”. La presenza di queste caratteristiche in più gruppi diversi fa pensare anche che il bipedismo sia nato indipendentemente in diversi momenti e in diversi gruppi.
Ci sono poi dei varani che riescono ad alzarsi su due zampe ma come fra i mammiferi le marmotte, si ergono stando fermi: questo comportamento sembra essere a scopo difensivo o per esplorare meglio i dintorni.
Il bipedismo delle lucertole attuali offre dei particolari apparentemente sconcertanti: non solo appare illogico che meno zampe ci sono più si corra, ma è stato persino dimostrato che non ci sono vantaggi né di velocità né di efficienza energetica nelle lucertole che corrono su due zampe [5] anche se le lucertole capaci di farlo, quando corrono su due zampe sono ai vertici come velocità di punta (e anche di lunghezza delle zampe posteriori, ma questo appare abbastanza ovvio...). Quanto poi al Basilisco, è ancora più "assurdo": distribuisce il peso su due zampe anziché su 4. Ma, allora, cosa ha portato al bipedismo? Banalmente, i vantaggi non stanno nella massima velocità o nel minore impiego di energia, ma nelle prestazioni migliori in fase di accelerazione e nella abilità di cambiare direzione più in fretta. Ed è accertato che queste lucertole lo fanno soltanto quando sono costrette a farlo da cause esterne.
 |
| Lo Stegosauro, quadrupede "di ritorno" già nel Giurassico inferiore |
LE MOTIVAZIONI DEL BIPEDISMO DEI DINOSAURI. I dinosauri triassici erano tutti bipedi obbligati (o quasi). Dal Giurassico in molti dinosauri erbivori l’aumento delle dimensioni e/o la formazione di armature hanno spesso determinato l’abbandono della stazione eretta, le cui tracce rimangono nella conformazione degli arti anteriori, più corti di quelli posteriori. Altri erbivori come gli iguanodontidi potevano alternare una andatura bipede, probabilmente più veloce, a quella quadrupede. Fra i teropodi solo lo spinosauro potrebbe aver usato entrambe le locomozioni. Il passaggio da una alimentazione carnivora ad una erbivora ha favorito il ritorno al quadrupedismo, a causa delle modifiche nella forma del corpo, in particolare della cavità addominale, dovute alla maggior complessità del tubo digerente di un erbivoro rispetto a quello di un carnivoro, che ne hanno spostato in avanti il baricentro.
Sulle motivazioni del bipedismo nei primi dinosauri ci sono diverse ipotesi:
- la termoregolazione e la possibilità di godere di una vista “dall’alto”: queste ipotesi erano nate quando era valido il “modello canguro”, secondo il quale i dinosauri stavano in piedi come i canguri. Oggi si ritiene che il corpo fosse in posizione suborizzontale come in molti uccelli: in questa posizione la testa non risulta più elevata rispetto al corpo, per cui queste ipotesi oggi sono poco fondate
- l’andatura bipede poteva servire per utilizzare meglio le zampe anteriori in funzione predatoria se non addirittura per consentire la manipolazione delle prede. La cosa è suggerita dal fatto che gli antenati dei dinosauri fossero carnivori ma ci sono forti dubbi sulle capacità cerebrali necessarie per questi compiti ed inoltre molti teropodi avevano arti anteriori piuttosto piccoli, sicuramente meno efficienti dei lunghi musi per questi scopi (anzi, questa diminuzione della lunghezza degli arti anteriori può addirittura essere interpretata come un segno della loro inutilità). Inoltre si tratta di un punto di vista un po' troppo antropocentrico, ma rimane una ipotesi piuttosto “gettonata”.
Una spiegazione brillante al proposito è stata appena presentata da W. Scott Persons e Philip J. Currie, paleontologi dell’Università dell’Alberta [6].
Per iniziare sono state esaminate le condizioni ecologiche in cui è iniziato il bipedismo dei dinosauri: i loro antenati erano piccoli carnivori che vivevano a contatto di predatori ben più massicci di cui potevano anche essere preda. Pertanto la loro vita doveva essere basata sull’agilità e sulla velocità, capacità di cui i loro predatori dalle zampe corte e dall’andatura strisciante erano scarsi.
È stata avanzata la possibilità che negli antenati dei dinosauri la capacità di mantenere questa andatura più a lungo, più dispendiosa dal punto di vista energetica almeno all’inizio, sia in qualche modo collegata ad un certo grado di omeotermia presente già oltre 200 milioni di anni fa.
Ho scritto che era più dispendiosa “almeno all’inizio”, perché è logico pensare che ci sia stato un passaggio da quadrupedi obbligati a bipedi obbligati in cui a poco a poco il bipedismo da occasionale è divenuto obbligatorio, probabilmente grazie alla modificazione di una serie di caratteristiche anatomiche, in particolare:
- l'irrobustimento di bacino, arti posteriori e coda per migliorare l’andatura bipede
- la riduzione degli arti anteriori che non essendo più usati per la locomozione, meno pesavano meglio era
Insomma, l’evoluzione, avrebbe favorito sempre di più in questo gruppo le forme capaci di passare più tempo possibile in stazione bipede con sempre maggiore velocità e agilità e minore dispendio energetico. Bisogna notare inoltre che se fra i sinapsidi del Permiano molti avevano una andatura strisciante (per esempio i pelicosauri), i terapsidi triassici (come i mammiferi attuali) tenevano il corpo sollevato. Ricordo anche che stare sollevati da terra permette una migliore ossigenazione e stressa meno gomiti e ginocchi.
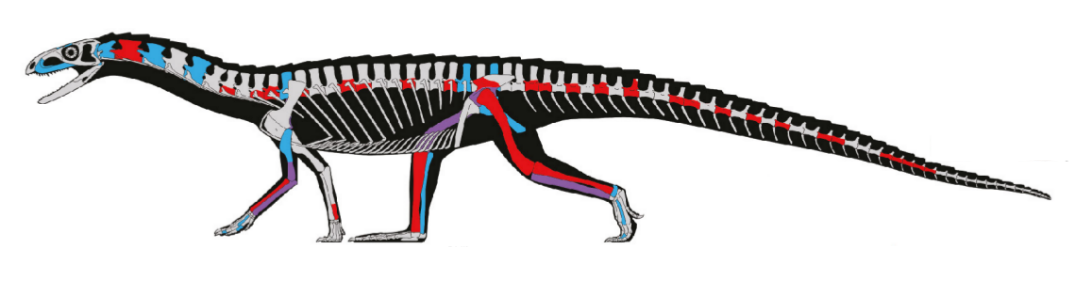
 |
| L'immagine mostra l'estensione del distretto muscolare caudofemorale in un tyrannosauro |
BIPEDISMO E DISTRETTO MUSCOLARE CAUDIFEMORALE. in tutti i modelli sulla locomozione dei dinosauri la coda svolge un ruolo fondamentale, con la sua massa in grado di spostare il baricentro del corpo verso la parte posteriore, scaricando dal peso la parte anteriore; di conseguenza per molti Autori le code lunghe e robuste degli arcosauri sono state un fattore essenziale per lo sviluppo della locomozione bipede. Le modifiche che hanno consentito una maggiore efficienza dell’andatura bipede dei dinosauri sono dovute essenzialmente all’irrobustimento della parte posteriore del corpo, nel distretto muscolare caudofemorale. Si tratta dei muscoli che collegano bacino e femore e servono a muovere la coda: data la sua posizione, questi muscoli hanno potuto ingrandirsi senza interferire particolarmente con altre strutture del corpo (l’unica cosa importante da realizzare era il conseguente irrobustimento delle estremità degli arti posteriori per sostenerne il peso).
Infatti l’andatura bipede, combinata con una muscolatura caudofemorale importante, comporta dei vantaggi nella corsa bipede perché questi muscoli riescono ad offrire delle possibilità maggiori di propulsione agli arti posteriori rispetto a quelli anteriori.
E che l’andatura bipede sia collegata alla corsa lo dimostra il fatto che solo forme erbivore dotate di doti difensive diverse da quelle della corsa sono ritornati dei quadrupedi obbligati: i grandi sauropodi, la cui massa troppo grande e la vita in branco impedivano la predazione (come oggi succede con gli elefanti) o quelli dotati di armature come i ceratopsidi: gli altri, anche se erano in grado di muoversi su 4 arti, come i canguri attuali se dovevano correre erano diventati obbligati a farlo su due zampe a causa della pesantezza maggiore della parte posteriore del corpo e del ridotto sviluppo degli arti anteriori.

MA PERCHÈ QUESTO NON È SUCCESSO CON I MAMMIFERI? Anche fra i mammiferi ci sono abbondanti esempi di animali che si basano sulla corsa per predare o per sopravvivere ai predatori. Ma nessuno di loro è un bipede preferenziale se non canguri che se devono andare veloci non possono che saltellare in andatura bipede, o obbligato come Homo: fra i Primati alcune specie camminano su due zampe per certi tratti e in più c’è qualche altro sporadico caso di bipedismo più o meno occasionale.
Questa differenza la troviamo anche nel Triassico. Ricordo che i rettili e gli uccelli sono diapsidi (hanno nel cranio due aperture temporali), mentre i mammiferi appartengono ai sinapsidi (una sola apertura temporale)..
Arcosauri e terapsidi sono stati nel Triassico i gruppi di vertebrati terrestri di vertice, differenziandosi in maniera molto vivace per occupare una gran quantità di nicchie ecologiche (quello che ora è riservato quasi esclusivamente ai mammiferi se si eccettuano le acque interne tropicali, dove i protagonisti sono sempre degli arcosauri (i coccodrilli). Eppure fra i sinapsidi del Triassico (compresi gli antenati dei mammiferi) non ci sono forme bipedi e fra i mammiferi mesozoici contiamo persino delle forme volanti (o quantomeno capaci di planare come Volaticotherium antiquus) ma nessun bipede.
Una differenza importante è la mancanza nei sinapsidi (almeno dal triassico in poi) del quarto trocantere, una struttura nel femore che serve appunto per l'inserzione di uno dei muscoli del distretto caudofemorale di cui invece sono ancora dotati i diapsidi.
Secondo Persons e Currie la chiave di questo sta nel sistema caudofemorale ridotto dei sinapsidi vissuti dal Triassico in poi. Nel Permiano anche molti sinapsidi avevano ben sviluppato questo sistema, ma sembra che i pochi sinapsidi sopravvissuti alla estinzione al passaggio con il Triassico siano stati esclusivamente delle forme che vivevano scavando tane nel sottosuolo. Per cui avevano perso nei tempi precedenti buona parte del sistema caudofemorale che in uno stile di vita come quello era un ingombro per diversi motivi:
- necessità di avere più sviluppate le zampe anteriori che scavano rispetto a quelle posteriori
- una coda lunga e robusta era di ostacolo per le manovre e i cambi di direzione nelle tane e inoltre in caso di pericolo quanto l’animale si rifugiarsi nella tana potrebbe esere agganciato dal predatore proprio dalla coda
La mancanza di un sistema caudofemorale importante è dunque la motivazione per cui i sinapsidi dal Triassico in poi, dai Terapsidi ai mammiferi, non hanno potuto a parte qualche caso particolare, sviluppare una andatura bipede.
I diapsidi che non hanno sviluppato andatura bipede non hanno avuto antenati per i quali l’agilità e l’accelerazione erano esigenze così preminenti.
[1] Berman, D.S., et al., 2000. Early Permian bipedal reptile. Science 290 (5493), 969–972.
[2] Ezcurra MD, Scheyer TM, Butler RJ (2014) The Origin and Early Evolution of Sauria: Reassessing the Permian Saurian Fossil Record and the Timing of the Crocodile – Lizard Divergence. PLoS ONE 9(2): e89165. doi:10.1371/journal.pone.0089165
[3] Brusatte, S.L., Niedźwiedzki, G., Butler, R.J., 2011. Footprints pull origin and
diversification of dinosaur stem lineage deep into Early Triassic. Proc. R. Soc. Lond. B: Biol. Sci. 278 (1708), 1107–1113.
[4] Nesbitt et al 2017 The earliest bird-line archosaurs and the assembly of the dinosaur body plan Nature, Aprll 2017 doi:10.1038/nature22037
[5] Clemente, C.J., Withers, P.C., Thompson, G., Lloyd, D., 2008. Why go bipedal? Locomotion and morphology in Australian agamid lizards. J. Exp. Biol. 211 (13),
2058–2065.
[6] Persons Currie 2017 The functional origin of dinosaur bipedalism: Cumulative evidence from bipedally inclined reptiles and disinclined mammals Journal of Theoretical Biology 420 1–7


Nessun commento:
Posta un commento